Alcune specie di Anolis presentano dimorfismo trofico, che può coinvolgere l’identità dell’oggetto (Rodríguez 2010), la quantità, la frequenza e il volume della preda (Perry 1996). In questi casi le differenze potrebbero essere correlate al dimorfismo delle dimensioni sessuali (dimensioni del corpo e dimensioni della testa) (Schoener 1968), al comportamento di foraggiamento (correlato alle differenze nei ruoli sessuali maschili e femminili) (Butler e Losos 2002, Butler et al., 2007, Perry 1996, Steffen 2009, Vincent e Herrel 2007), variazione nell’uso del pesce persico (Perry 1996, Schoener 1967), densità di popolazione (Francobolli et al. 1997), o differenze di fabbisogno energetico durante la stagione riproduttiva (Schoener 1968, Vitt e Zani 1998).
Le diete degli anoli variano tra e all’interno delle specie, e almeno alcune di queste variazioni sembrano legate alle differenze nell’uso delle risorse strutturali (tipo e altezza del pesce persico) e al relativo accesso e abbondanza di prede diverse (Fleming and Hooker 1975, Vitt and Zani 1998, Vitt et al. 2002, 2003b)., All’interno delle specie la variazione nell’ecologia alimentare è stata registrata in anoli amazzonici come A. trachyderma (Vitt et al. 2002), A. punctatus e A. transversalis (Vitt et al. 2003a), e A. fuscoauratus (Vitt et al. 2003b).
La variazione geografica di fattori quali interazioni interspecifiche competitive, disponibilità di prede e uso delle risorse strutturali è stata suggerita come causa alla base della variazione intraspecifica nelle abitudini alimentari in queste specie (Fleming e Hooker 1975, Gutiérrez e Rumiz 2002, Régnière 2009, Rodríguez 2010, Schoener 1968) e lucertole in generale (Aun et al. 1999).,
Anolis ventrimaculatus Boulenger, 1911 è una specie andina diffusa in tutta la Colombia occidentale. Questa specie mostra dimorfismo sessuale e forma del corpo, che varia anche tra le località (Calderón-Espinosa et al. 2013). Il dimorfismo in questi tratti è indicativo di differenze nell’uso delle risorse tra i sessi e possibilmente, almeno qualche variazione tra le popolazioni. Abbiamo valutato questa ipotesi descrivendo le diete di maschi e femmine in diverse località all’interno della gamma delle specie.,
MATERIALI E METODI
Abbiamo misurato la lunghezza dello sfiato del muso (SVL) di ogni individuo con un calibro digitale a 0,1 mm e rimosso lo stomaco e le porzioni prossimali dell’intestino. Stomaci e contenuto intestinale sono stati identificati per ordine e famiglia quando possibile, dopo Triplehorn e Johnson (2005). Abbiamo quindi stimato il numero di prede utilizzando teste, ali, zampe, antenne e altre parti del corpo identificabili. Poiché le prede sono state parzialmente digerite, non abbiamo potuto stimare le dimensioni delle prede. Successivamente in questo articolo, “articolo” si riferisce all’identità tassonomica (ad es.,, Ortotteri, Coleotteri), mentre “preda” si riferisce a singole prede ingerite dalle lucertole.
Analisi dei dati. Abbiamo valutato il dimorfismo alimentare a livello di specie e località confrontando il numero e la frequenza degli elementi osservati (quelli identificati a livello di ordine) tra i sessi utilizzando test Chi square. Il numero di prede è stato confrontato con i test t, dopo aver valutato qualsiasi relazione con lizard SVL utilizzando i test di correlazione Spearman (Zaar 1990). Abbiamo anche analizzato le percentuali di utilizzo di elementi che rappresentavano > 70% di ingesta, e utilizzato t test per confrontarli tra i sessi., L’uso delle prede è stato derivato dal numero di prede per oggetto per individuo/numero totale di prede ingerite da ciascun individuo.
Abbiamo anche analizzato la variazione geografica utilizzando ANOVA e a posteriori Tukey test per confrontare le diete di femmine e maschi e la percentuale di utilizzo di articoli che rappresentavano> il 70% di ingesta nelle varie località.
La normalità e l’omocedasticità sono state valutate per tutti i dati e trasformate in Log10 quelle che non soddisfacevano le ipotesi. Le analisi dei dati sono state eseguite in Statistica ver. 8.0 (demo).,
Ampiezza di nicchia trofica, sovrapposizione di nicchia e indici di dominanza. Abbiamo stimato la nicchia trofica in largo per i maschi e per le femmine, utilizzando l’inverso indice di Simpson (1949), come segue: B = (Σpi2)-1,
dove p rappresenta la percentuale di utilizzo della voce che ho. Abbiamo determinato anche se i maschi e le femmine simile risorse alimentari, stimando nicchia si sovrappongono, come proposto da Pianka (1973):
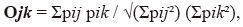
dove pij pik rappresenta la percentuale di utilizzo della risorsa che ho da femmine (j) e maschi (k). Questi indici sono stati calcolati utilizzando il software Ecosim.,
Infine, abbiamo stimato l’indice Lambda dominance (Aun et al. 1999, Montori 1988, 1991, 1992) come segue:
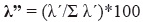
In questa equazione, λ’ = λ/n*100 e λ = Σpi2,
RISULTATI
Tutti i 118 individui esaminati avevano alcuni ingesta nello stomaco e nell’intestino prossimale. Un totale di 1021 prede, (593 nelle femmine e 428 nei maschi) sono state classificate in 16 articoli a livello ordinale di insetti, aracnidi (Araneae), miriapodi e crostacei (gli isopodi erano presenti in una femmina)., Imenotteri e coleotteri, che mostravano la più alta diversità tassonomica a livello familiare, sono stati classificati in 24 famiglie. Alcuni individui avevano ingerito materiale vegetale identificato come Marchantiophyta, shed e piccole pietre. Questi sono stati considerati incidentali, perché sono stati osservati solo in tre individui (tabella 1) e presumibilmente sono stati ingeriti avventatamente.,
Queste lucertole si nutrono principalmente di coleotteri, Imenotteri (principalmente formiche) e larve di insetti, con coleotteri classificati come elemento primario, mentre Imenotteri e larve sono stati classificati come elementi secondari, e altri sono stati considerati accidentali (tabella 1). La maggior parte degli stomaci di tutte le località conteneva nematodi parassiti non identificati.
Dimorfismo sessuale e dieta. Quattordici articoli sono stati osservati nello stomaco di ogni sesso. Maschi e femmine si erano nutriti con lo stesso numero di articoli (X2 = 1.37, p > 0.05). Tuttavia, alcuni elementi sono stati osservati solo nei maschi o nelle femmine., Isopodi e isopterani (prede piccole e per lo più terrestri) sono stati trovati solo in una e due femmine, rispettivamente, mentre trichopterani e neuropterani (prede grandi e per lo più arboricole) erano presenti solo in due maschi provenienti da diverse località. Nonostante queste differenze, le diete maschili e femminili erano molto simili, come indicato dalla sovrapposizione di nicchia molto alta (0.99). Le larghezze di nicchia erano 6,09 per i maschi e 6,17 per le femmine.
I maschi e le femmine della località hanno mostrato una diversa percentuale di utilizzo di ortotteri (t = 3,76, p = 0,002) e imenotteri (t = 2,44, p = 0.,02), e quelli della località sei hanno ingerito diverse quantità di ortotteri (t = 2.24, p = 0.04). Nella località due, solo le femmine si nutrivano di ortotteri. Maschi e femmine in altre località non differivano significativamente nell’uso di questi elementi.
Variazione geografica. Solo quegli articoli che erano apparentemente esclusivi di maschi o femmine, e presenti in frequenze molto basse e basse percentuali di utilizzo mostravano variazioni geografiche., Gli isopodi sono stati osservati solo nella località uno, gli isopterani sono stati trovati in due femmine nella località sei, trichopterani nella località tre, neuropterani nella località due, Blattodea nelle località due e sei, miriapodi nelle località tre e sei e collembolani nelle località tre e quattro (figura 1).
I maschi mangiavano un numero simile di prede in tutte le località, ma le femmine no (F60, 5 = 3,56, p = 0,007) con differenze significative osservate tra le femmine della località uno e quelle delle località tre, quattro e cinque (Tukey test, p< 0,05).,
DISCUSSIONE
Anolis ventrimaculatus è un insettivoro come la maggior parte delle altre specie di Anolis tra cui A. tolimensis (Ardila-Marín et al. 2008), A. homolechis (Rodríguez 2010), A. cybotes (Fobes et al. 1992), e A. aeneus (Francobolli et al. 1997). Coleotteri, imenotteri, e larve di insetti erano gli elementi più frequentemente consumati, probabilmente perché questi elementi sono più abbondanti (Fobes et al. 1992). I nostri risultati suggeriscono anche che questa specie può essere considerata una dieta generalista / opportunista che foraggia principalmente nei posatoi degli alberi e raramente sul terreno.,
I nematodi trovati nello stomaco e nel contenuto intestinale sono parassiti comuni nelle lucertole di Anolis (Fobes et al. 1992, Goldberg e Bursey 2002, Lenart et al. 1994). Il carico parassitario potrebbe influenzare il successo riproduttivo e il comportamento della lucertola (Suzán-Azpiril et al. 2008), ma tale informazione non è disponibile per A. ventrimaculatus.
Le differenze sessuali nella dieta erano minime ed erano principalmente correlate al numero di prede, con i maschi che probabilmente mangiavano prede più grandi, spiegando il minor numero di prede ingerite rispetto alle femmine e come osservato in altri anoli (ad es.,, Perry 1996, Rodríguez 2010, Schoener 1967, 1968). Anche se non formalmente determinato a causa della digestione parziale di ingesta, differenze nelle dimensioni delle prede sono state suggerite da alcuni oggetti molto grandi trovati solo nei maschi (Montealegre 1997, Triplehorn e Johnson 2005).
Le differenze minori osservate tra i sessi sono simili a quelle osservate in A. trachyderma (Vitt et al. 2002) e A. eneus e A. richardii (Simmons et al. 2005)., Alcune delle differenze, come il numero di prede o la percentuale di utilizzo dell’oggetto, potrebbero essere correlate a differenze nella disponibilità di prede attribuibili a diversi usi strutturali di microhabitat tra i sessi, come suggerito dalla variazione delle dimensioni del corpo e del dimorfismo della forma (Calderón-Espinosa et al. 2013). Tuttavia, l’uso di microhabitat o la disponibilità di prede in questa specie non è stata descritta e le cause che sono alla base delle differenze sessuali minori nella dieta di A. ventrimaculatus rimangono sconosciute.,
La variazione geografica osservata nell’identità dell’elemento all’interno dei maschi e delle femmine potrebbe essere spiegata come un effetto di dimensione del campione, poiché questa variazione riguardava solo quegli elementi, trovati a frequenze molto basse in ogni località. Questi elementi sono stati osservati nella dieta di altre specie a frequenze altrettanto basse (Ardila-Marín et al. 2008, Rodríguez 2010).
Il numero totale di prede variava solo tra le femmine di diverse località., Il consumo di quantità di cibo potrebbe essere correlato alla condizione riproduttiva degli individui, poiché i requisiti energetici delle femmine gravide sono diversi da quelli delle femmine non gravide (Andrews e Asato 1977, Losos 2009, Rodríguez 2010). Tuttavia, la maggior parte delle femmine incluse in questo studio erano gravide, solo tre erano in uno stadio vitellogenico precoce e una era postgravida. Di conseguenza, la variazione della quantità di cibo ingerito dalle femmine non era apparentemente correlata al fabbisogno energetico per lo sviluppo del follicolo o dell’uovo.,
Inoltre, le fluttuazioni stagionali nella disponibilità di prede potrebbero influenzare le abitudini alimentari di queste lucertole. Le densità di insetti e altri invertebrati variano solitamente tra le stagioni (Régnière 2009), influenzando la disponibilità di prede per le lucertole, come descritto per A. homolequis (Rodríguez 2010), A. cupreus (Fleming and Hooker 1975) e altre specie di questo gruppo (Aun et al. 1999).
Il nostro studio rappresenta un primo sforzo per comprendere gli effetti del dimorfismo delle dimensioni del corpo e della forma e la variazione geografica in un anolo andino., Tuttavia, sono necessari ulteriori dati ecologici per spiegare la variazione alimentare minore osservata in questa specie.
AKKNOWLEDGEMENTS
Questo studio è stato reso possibile attraverso l’accesso a campioni conservati conservati nella collezione di rettili dell’Instituto de Ciencias Naturales, ONU. Ringraziamo C. Sarmiento, G. Amat, E. Flores, J. H. García, D. Martínez e F. Fernández per l’aiuto nell’identificazione degli artropodi e J. Uribe aiutando a identificare il contenuto vegetale degli stomaci di lucertola. Una recensione anonima ha migliorato questo manoscritto.
Andrews RM, Asato T. 1977., Utilizzo energetico di una lucertola tropicale. Biochimica comparata e fisiologia, 58A: 57-62.
Boulenger GA. 1911. Descrizioni di nuovi rettili delle Ande del Sud America, conservati nel British Museum. Gli annali e la rivista di Storia naturale, 8a serie, 7 (37): 19-25.
Butler MA, Losos JB. 2002. Dimorfismo sessuale multivariato, selezione sessuale e adattamento nelle lucertole Anolis delle Grandi Antille. Monografie ecologiche, 72 (4): 541- 559.
Calderón-Espinosa ML, Ortega-León A, Zamora-Abrego JG. 2013., Variazione intraspecifica delle dimensioni corporee in una specie di anolo dell’altopiano andino (Anolis ventrimaculatus: Squamata, Dactyloidae). Revista de Biología Tropical, 61 (1): 255-262.
Eaton JM, Larimer SC, Howard KG, Powell R Parmerlee JS Jr.2002. Densità di popolazione e rilascio ecologico della lucertola solitaria Anolis gingivinus in Anguilla, Indie occidentali. Caribbean Journal of Science, 38 (1): 27-36.
Gutiérrez T, Rumiz D. 2002. Patrones de diversidad de grupos selectos de insettos en el bosque chiquitano y pampas del cerrado de Santiago y Tucavaca, Santa Cruz, Bolivia., Revista Boliviana de Ecología, 11 (1): 37-46.
Henderson EW, Powell R. 2009. Storia naturale dei rettili e degli anfibi dell’India occidentale. Gainesville (Florida): Università della Florida Press. pag. 495.
Lenart LA, Powell R, Parmelee JS Jr, Smith DD, Lathrop A. 1994. Dieta e un parassita gastrico di Anolis armouri, un anolo cybotoide dalle foreste di pini montane nel sud di Hispaniola. Storia naturale erpetologica, 2 (1): 97-100.
Perry G. 1996., L’evoluzione del dimorfismo sessuale nella lucertola Anolis polylepis( Iguania): evidenza dalla variazione intraspecifica nel comportamento di foraggiamento e nella dieta. Canadian Journal of Zoology, 74 (7): 1238-1245.
Pianka ER. 1973. La struttura delle comunità di lucertole. Revisione annuale di ecologia e sistematica, 4: 53-74.
Régnière J. 2009. Predicción de la distribución continental de insectos a partir de la fisiología de las especies. Unasilva, 60 (1): 231-232.
Schoener TW. 1967. Il significato ecologico del dimorfismo sessuale in termini di dimensioni nella lucertola Anolis conspersus., Scienza, 155 (3761): 474-477.
Simpson EH. 1949. Misurazione della diversità. Natura, 163: 688.
Francobolli JA, Losos JB, Andrews RM. 1997. Uno studio comparativo della densità di popolazione e dimorfismo delle dimensioni sessuali nelle lucertole. Il naturalista americano, 149 (1): 126-135.
Steffen JE. 2009. Una valutazione dell’allometria per il dimorfismo delle dimensioni sessuali negli anoli continentali. Rivista sudamericana di Erpetologia, 4 (3): 245-252.
Suzán-Azpiril G, Galindo-Maldonado F, Ceballos-González G. 2008. La importancia del estudio de enfermedades en la conservación de la fauna silvestre., Veterinaria México, 31 (3): 223-230.
Vincent SE, Herrel A. 2007. Correlati funzionali ed ecologici di dimorfismi a base ecologica nei rettili squamati. Biologia integrativa e comparativa, 47 (2): 172-188.
Vitt LJ, Zani PA. 1998. Uso preda tra specie lucertola simpatrica in pianura foresta pluviale del Nicaragua. Journal of Tropical Ecology, 14 (4): 537-559.
Vitt LJ, Avila T, Zani PA, Espósito C. 2002. La vita in ombra: L’ecologia di Anolis trachyderma (Squamata: Polychrotidae) in Amazzonia Ecuador e Brasile, con confronti con anole ecologicamente simili., Copeia, 2002 (2): 275-286.
Vitt LJ, Avila T, Zani PA, Sartorius S, Esposito C. 2003b. Life above ground: Ecology of Anolis fuscoauratus in the Amazon rain forest, and comparations with its nearest relatives. Canadian Journal of Zoology, 81 (1): 142-156.
Zaar JH. 1990. Analisi biostatistica. 4a ed. Upper Saddle River, Nueva Jersey: Prentice-Sala stampa. pag. 663.