Introducere
Secvențe de alela membre sau „motive”, bazat pe polimorfisme la genomul mitocondrial (mtDNA) și non-recombinarea porțiune a cromozomului Y (NRY) au fost propuse pentru a urmări Evreu popoare. În special, Skorecki și colab., (1997) a sugerat mai întâi, că diferența de distribuție a Y-ADN haplotipurilor bazate pe doi markeri, Y Alu polimorfism (YAP) și Y-STR (Short Tandem Repeat) DYS19, ar putea fi un proxy de patrilineal coborâre de Cohanim mari preoți. Haplotipul Yap – / DYS19 B a fost recunoscut ca posibil haplotip fondator al Preoției evreiești. La scurt timp după aceea, Thomas și colab. (1998) a rafinat această ipoteză pe baza variabilității la 6 y-SNPs (polimorfisme cu un singur nucleotid) și 6 y-STRs., Un haplotip modal a fost găsit în cromozomii Cohen Y împreună cu un grup de haplotipuri strâns legate, indiferent dacă aparțineau comunităților Ashkenazim sau Sefardice, a căror coalescență a fost datată despre timpul Regatului David (acum 2619-3221 ani). Nebel și colab. (2000, 2001) a definit Cohen Modal Haplotip sau CMH (Tabelul 1) ca o 6-locus Y-STR haplotip aparținând haplogrupul Ue10, care este J1-M267 în conformitate cu nomenclatura actuală (Y-ADN Haplogrupul Copac 2014 Versiune: 9.70)., Astfel de motiv a dus la 2-3 mutatii câțiva pași de alte Ue10 modal haplotipuri observate în arabă vorbitoare de grupuri (MH Galileea, MH Beduin, MH Palestinieni) și la un pas de paraphyletic modal haplotip Musulmane Kurzi care aparțin haplogrupul Ue9 (J2-M172 în nomenclatura actuală). Într-un studiu ulterior, Thomas si colegii sai au raportat motive evreiești CMH, de asemenea, dintr-un set de haplotipuri din Africa de Sud-Est (Thomas et al., 2000).în cel mai recent studiu pe această temă, Hammer et al., (2009) au încercat să investigheze în continuare Y-ADN dovezi biblice coborâre de Cohanim dintr-un singur strămoș (biblic Aaron) prin extinderea discriminare puterea de CMH la 6 – la 12-locus Y-STR haplotipuri (eCMH, Tabelul 1). Ei au susținut că originea diversității asociate cromozomilor Cohanim J1-P58 ar putea fi datată între 4280 și 2100 de ani în urmă.

Tabelul 1. NRY haplotype motive.,haplotipurile Levite modale (LMH și eLMH, Tabelul 1) au fost în schimb propuse de behar et al. (2003) ca haplotipuri modale alternative Y-STR în cadrul variației haplogrupului R1a-M17 a Leviților Ashkenazim, parțial împărtășită cu est-europenii. Această constatare a declanșat ipoteza originii Idiș dintr-o limbă Slavă (Sorbian) și introgresia Khazarian cromozomi Y în formarea inițială de Evrei Ashkenazi 1000 de ani în urmă., Cu toate acestea, resuscitare analize găsit diverse fondator evenimente, printre Ashkenazi Leviți în R1a care să demonstreze că un anumit sub-grup, la M582, ar fi o semnătură de Aproape de origine Est în pre-Diaspora ori (Rootsi et al., 2013).
Thomas și colab. (2002) a găsit semnale de evenimente fondatoare mult mai puternice în liniile specifice femeilor (mtDNA) din diferite comunități evreiești decât în liniile corespunzătoare specifice bărbaților., Ei au găsit cel puțin 8 haplotipuri modale (frecvență >10%) în regiunea HVS-I a ADN-ului mitocondrial în 8 comunități de evrei separate geografic, în timp ce nu s-a observat nicio diferențiere la haplotipurile ADN-Y ale populațiilor evreiești și gazdă.
Folosind o înaltă rezoluție analiza haplogroups K și N, Behar și colaboratorii săi au identificat patru mtDNA fondator linii ca linie maternă sursă de aproximativ 40% din populația actuală de Evrei Ashkenazi (Behar et al., 2006)., Aceste linii au fost descrise ca fiind originare din Orientul Mijlociu în urmă cu aproximativ 2100 de ani „probabil dintr-o piscină mtDNA Ebraică/levantină.”Cu toate acestea, originea matrilineală complexă a Ashkenazi pare să fi fost cel mai bine reprezentată de Costa et al. (2013), OMS a dezvăluit că marea majoritate a liniilor materne Ashkenazi au fost rezultatul unor evenimente de adiție reiterate în Europa. În ceea ce privește potențialii confounders demografice ale scenariului de mai sus, Behar și colab. (2004) și Guha și colab., (2012) au susținut o derivă genetică puternică pentru a contribui la frecvența neobișnuit de ridicată a alelelor bolii recesive și a diversității scăzute a mtDNA și a ADN-ului Y în populațiile Ashkenazi. Pe de altă parte, literatura recentă bazată pe analize la nivelul genomului (Atzmon et al., 2010; Behar și colab., 2010; Bray și colab., 2010) a evidențiat rolul decisiv al amestecului în conturarea actualului bazin de ADN Evreiesc.în ultimii ani, companiile genealogice genetice au recrutat zeci de mii de voluntari care au acceptat să împărtășească rezultatele testelor lor genetice în depozite publice., Cea mai mare bază de date de strămoși disponibilă până în prezent este Family Tree ADN archive (FtDNA), incluzând mai mult de 506 k înregistrări pentru Y-ADN și aproximativ 180 k înregistrări pentru mtDNA împărțite în aproape 8 K proiecte, unde participanții își pot împărtăși propriul profil ADN pentru a urmări o moștenire comună după nume, descendență sau geografie. Folosind o astfel de resursă valoroasă, am construit o bază de date vastă de profiluri mtDNA și Y-ADN disponibile din >600 proiecte FtDNA., Am explorat baza de date pentru a studia variabilitatea asociată motivelor genetice propuse de literatura de specialitate în voluntari care pretind o origine evreiască și în grupuri non-evreiești de origine europeană, africană sau Orientul Mijlociu.
Noi menite să ofere o actualizare experimentale de fond prin care să argumentul defectele și capcane pot apărea atunci când se utilizează haplotip motive ca diagnostic markeri de origine Evreiască.,
materiale și metode
construirea bazelor de date
haplotipurile Y-STR și Y-SNP-urile au fost descărcate din arborele genealogic ADN browser-ul cromozomului Y (https://www.familytreedna.com/projects.aspx). Pentru a se potrivi motive de referință (Behar et al., 2003) cu liniile directoare ale Nomenclaturii standard (Gusmão et al., 2006) stările de alelă la DYS439 au fost corectate scăzând 4 repetări. Haplotipurile HVS-I și SNP-urile mtDNA au fost descărcate din browserul de cromozomi genealogici ADN mt (https://www.familytreedna.com/projects.aspx).,înregistrările (https://www.familytreedna.com/projects.aspx) au fost mai întâi împărțite în proiecte care vizează în mod explicit reconstruirea strămoșilor evrei și a altor proiecte. În al doilea rând, am filtrat în cadrul și printre proiecte acele înregistrări cu numere de kit duplicate și origini incerte. Atunci când nu sunt disponibile, asociațiile dintre haplotipuri și SNP-uri s-au făcut după numărul kitului., În cele din urmă, am scos haplotipuri care nu se potrivesc un dublu criteriu: NRY-haplotipuri – a fi dactilografiate la un minim de 12 Y-STR loci (set de „extins” motive) și atribuite haplogroups fie prin direct SNP dactilografiere sau de la previziuni pe baza ≥25-locus STR profile; mtDNA-haplotipuri—să fie secvențiat cel puțin pentru HVS-am site-uri 16,024–16.569 de și atribuite haplogroups prin tastarea directă de diagnostic Snp. Am obținut o grilă finală de 62,920 înregistrări y-ADN și 30,469 înregistrări mtDNA. Bazele de date au fost căutate pentru 6 și 12 motive Y-STR (evreiești și arabe) și pentru 4 motive HVS-i Ashkenazi (tabele 1, 2).,

Tabelul 2. mtDNA haplotype motive.o limitare a arhivelor genealogice genetice disponibile publicului este auto-atribuirea participanților la categorii sociale / etnice, cum este cazul descendenței evreiești. O alta este presupusa relație dintre participanții din cadrul aceluiași grup etnic., În timp ce fostul limita este greu să se stabilească, și surse de eroare, cum ar fi adopțiile și nelegitim paternities nu poate fi exclusă, printr-o analiză preliminară strânsă înrudire pare a afecta doar marginal statistici sumare prezentate în această lucrare cu privire la cele anterioare, rapoarte științifice. Înregistrările care aveau un strămoș comun au fost 12 din 3903 cunoscuți (0,3%) în proiectele de strămoși evrei și 606 din 48,006 cunoscuți (1,5%) în celelalte proiecte.,
Calculul Haplotip Ratele de Mutatie
Am utilizat următoarele pedigree pe bază de tarife:
Medie mutație ratele pe Y-haplotip au fost obținute în conformitate cu „genealogice” metoda (Klyosov, 2009a; Rozhanskii și Klyosov, 2011) după calibrare pentru mutațiile descrise în acesta, și să „germ-line” estimările combinarea de la tată la fiu perechi de date din literatura de specialitate (29 de studii revizuite de către Burgarella și Navascue e, 2011, plus Ballantyne et al., 2010).,
În primul caz panta dreptei de calibrare complot pentru 12-locus Rts de eCMH și eLMH motive se potrivește bine la o valoare de rata de mutație constantă de 0.00166 mutație pe marker per condiționată generație de 25 de ani, care este 0.020 mutații pe haplotip la fiecare 25 de ani.
În acest din urmă caz, numărul de observat mutații într-un total de 126,873 meioses (Tabelul S1) a dat o rată medie 2.113 × 10-3 (± 1.369 × 10-3) mutații pe marker per generație, care este de 0.025 ± 0.016 mutații pe haplotip per generație., Pentru trimerică locus DYS426, în cazul în care nu au observat mutații peste pedigree, o regresie rata bazează pe o logistică populației model a fost folosit (0.458 × 10-3 mut/gen, Burgarella și Navascue lui, 2011).ca rate medii de mutație pe haplotip HVSI am adoptat rata cumulată bazată pe pedigree în Howell et al. (2003) pe baza a 11 studii din literatura de specialitate: 1,06 × 10-2 mut/gen presupunând un interval de generație de 25 de ani. Este de aproximativ șapte ori mai mare decât rata calibrată fosil pentru segmentul 16,051–16,400 după Soares și colab. (2009): 1.,42 × 10-3 mut / gen presupunând un interval de generație de 25 de ani.
Rețea Analize
utilizarea rețelelor avut un dublu scop: multe dintre între-haplotipuri intermediar mutatii pași, eventual acoperit de recurente mutație sau prin incomplete de eșantionare, ar fi putut fi recuperat și luate în considerare în estimări de timp; poziția de haplotip motive pe cel mai bun copac dat un indiciu lor filogenie.,
Mutatii relațiile între Y-ADN-ul 12-locus motive și HVS-I (16,024–16.569 de bp) motive de la persoane fizice de auto-raportate de origine Evreiască au fost vizualizate prin intermediul mediană alături de rețea algoritm implementat în Rețeaua de 4.612 software (http://www.fluxus-engineering.com) potrivit Zalloua et al. (2008). La construirea rețelelor, valoarea implicită (10) a fost dată fiecărui sit HVS-I și un scor calculat pe varianța estimată la fiecare locus a fost adoptat pentru datele Y-STR (scor pentru un locus dat = 10 * varianța totală peste toate loci/varianța la acel locus dat).,
Pentru Y-ADN-ul, am selectat doar haplotipuri (N = 142) atribuite R1a1a1-M417 neam și în aval subclades conține LMH motiv (16-12-25-10-11-13, Tabelul 1), și haplotipuri (N = 73) atribuite J1a2b-P58 neam și în aval subclades conține CMH motiv (14-16-23-10-11-12, Tabelul 1). Locus DYS385 nu a fost folosit deoarece alele nu pot fi atribuite corect specifice duplicat regiune, și DYS389II a fost tratată ca DYS389b = DYS389II – DYS389I., Timpii de coalescență au fost estimați din rețele de către statistica rho folosind rate bazate pe „pedigree”- precum și „genealogic”.
Pentru mtDNA, am selectat doar haplotipuri care conțin cele trei motive K (16093C 16224C 16311C 16519C 16524G, motif_K1; 16224C 16234T 16311C 16519C, motif_K2; 16224C 16311C 16519C, motif_K3; Tabelul 2). Timpii de coalescență au fost estimați din rețele de către statistica rho folosind rate bazate pe „pedigree”.,teorema Bayes a fost utilizată pentru a calcula probabilitatea condiționată a unui individ de a aparține populației evreiești, având în vedere că astfel de indivizi poartă un motiv „diagnostic” NRY sau mtDNA. O astfel de probabilitate este funcția frecvenței unui haplotip dat la evrei și non-evrei (obținută din tabelele 3-5) și proporția indivizilor evrei (estimată a fi de ordinul a 13 milioane) în populația globală (7, 2 miliarde de oameni).

Tabelul 3., Distribuția NRY (tastat și prezis pe Y-STRs) și motive mtDNA în baza de date publică FTDNA.
rezultate
analiza înregistrărilor FtDNA (tabelele 3-5) a confirmat că niciun motiv genetic transmis de-a lungul liniei materne sau paterne nu este exclusiv pentru evrei. Cu toate acestea, nu a fost posibilă extinderea acestei concluzii la subgrupuri evreiești, cum ar fi Leviții și Cohanim, deoarece acest statut este rareori auto-raportat în intrările FtDNA., Singura excepție este cromozomul Y „proiectul Cohen Zadokites” care se alătură descendenților presupuși ai nepotului lui Aaron Zadok. Acolo, într-adevăr, am putea verifica (tabelul S2) că participanții aparțineau unui total de 6 haplogrupuri diferite cu J2, nu J1, ca modal (63%). Haplotipul No 12-locus a fost observat ca fiind privat pentru membrii proiectului, iar J1-eCMH a însumat doar 4 din cele 59 de înregistrări cu origine cunoscută (~8%).,
analiza Y haplotip distribuții în parental crengile copacilor a confirmat că, deși nu există o separare clară între distribuția de CMH, eCMH, LMH, și eLMH între Evrei și non-Evrei populații (chi pătrat rezultatele în Tabelul 4), niciun motiv este diagnostic de monofiletic haplogroups când probabilitatea condiționată de cesiune este estimată folosind formula Bayes (Figura 1). Chiar și cele mai rezolvat Y-ADN motive (eLMH și eCMH) au fost găsite în două sau mai independent haplogroups a cărei parte superioară ori de divergență sunt estimate la nu mai puțin de 30 de K ani., Acest lucru ar putea fi explicat printr-o astfel de adânc originea și evoluția ulterioară, fără nici o schimbare, de efectele secundare ale nu a recunoscut paternitatea, gene de conversie sau, cel mai probabil, având în vedere rata de haplotip schimba (aproximativ 24 de mutații sunt de așteptat să apară în 30 de K ani la extended Y-STR haplotipuri) și frecvența mare a polyphyletism, de homoplasy.

Tabelul 4. Distribuția motivelor în haplogrupurile Y-ADN (Hg) între 5281 evrei (J) și 57,639 non-evrei (NJ).,

Figura 1. Estimarea puterii de diagnosticare a unui set de haplotipuri modale pentru a atribui un eșantion dat populației evreiești. Fiecare linie arată probabilitatea condiționată (Formula Bayes) de a atribui un anumit individ care poartă haplotipuri modale mtDNA (A) sau Ychr (B) populației evreiești în funcție de proporția indivizilor evrei din eșantion. Punctele roșii arată probabilitatea posterioară obținută la introducerea fracției evreiești estimate a populației mondiale.,
este de remarcat faptul că pozițiile de-a lungul copaci de eLMH (central, Figura 2A) și eCMH (periferice, Figura 2B) sugerează că acesta din urmă s-ar putea nu au fost prezente în primele piscină de fondatori, ci pur și simplu să fie rezultatul unei mai recentă expansiune.

Figura 2. Unul dintre cei mai parcimonioși trei copaci construiți de algoritmul de rețea Median-Joining., Am folosit 12-locus Y-STR haplotipuri conține LMH motiv atribuite R1a-M417 lineage (A) și CMH motiv atribuite J1-P58 lineage (B), împreună cu lor din aval subclades în indivizi care pretind că o origine Evreiască. Cercurile reprezintă haplotipuri, cu zone proporționale cu numărul de indivizi pe care îi conțin.analiza distribuțiilor haplotipului mitocondrial la evrei indică faptul că, cel puțin cu nivelul actual de rezoluție SNP, un singur motiv (K3) din patru a fost găsit în haplogrupuri diferite (Tabelul 5).,
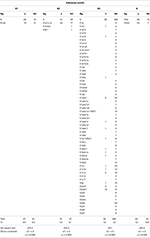
Tabelul 5. Distribuția celor patru motive Ashkenazi mtDNA în haplogrupuri (Hg) între 2818 evrei (J) și 27,651 non-evrei (NJ).pe de altă parte, constatările de mai sus au subliniat o supra-reprezentare a acestor motive la evrei în comparație cu non-evrei, precum și o acumulare semnificativă de motive în anumite haplogrupuri (tabelele 4, 5)., R1a Evreu haplotipuri care transportă Ashkenazi Levit motiv LMH (Figura 2A) par să împărtășească CTS6 variantă, a cărui TMRCA a fost estimată a fi între 1175 ± 341 de ani în urmă (folosind genealogice rata) și 924 ± 268 de ani în urmă (folosind germeni-rata de linie). De asemenea, J1 Evreu haplotipuri afișarea Cohanim motiv CMH (Figura 2B) par să împărtășească YSC0000234 variantă, a cărui TMRCA a fost estimată a fi între 1255 ± 441 de ani în urmă (folosind genealogice rata) și 986 ± 346 de ani în urmă (folosind germeni-rata de linie)., Evrei haplotipuri care transportă K1, K2 și N motive sunt exclusive, respectiv, de K1a9, K1a1b1a, și N1b haplogroups (Tabelul 2). Divergență ori calculat la variabilitatea observată la aceste haplogroups plus K2a2a1 haplogroups dat recente date pentru stramosul comun al tuturor mtDNA motive, cu cele pentru K2 și K3, respectiv, 1370 ± 1241 ani în urmă și 1265 ± 639 de ani în urmă, comparabile cu ori dedusă pentru strămoși comuni extins Y haplotipuri.,
discuție
motivele genetice realizate din seturi de markeri haploizi non-recombinanți au fost folosite de mult timp pentru a urmări originile evreiești presupuse ale indivizilor singuri sau ale populațiilor întregi. Astfel de motive uni-parentale au fost, de asemenea, propuse pentru a atribui strămoși în studiile de asociere și în cazurile medico-legale.eșecul acestei practici este inerent naturii variației genetice. Ca o estimare conservatoare ne putem aștepta la o mutație nouă la fiecare 94 de generații (28 de substituții la fiecare 2633 de transmisii, Howell et al.,, 2003) în liniile mitocondriale și aproximativ la fiecare trei generații (3 × 10-8 mut/site/gen în pedigreele cu înrădăcinare profundă peste 10.15 Mb, Xue et al., 2009) în liniile NRY.prin urmare, în astfel de sisteme genetice, cu cât numărul de markeri tipizați este mai mare, cu atât este mai mică probabilitatea de a afla seturi de alele care se potrivesc exact cu secvența strămoșilor fondatori., Mai precis, o mutație ar avea loc la fiecare 40-50 de transmisii la extended Y-STR haplotipuri, fiecare 80-100 de transmisii de la clasic 6-locus Y-STR haplotipuri și fiecare 140-150 transmisii la 16,024–16.569 de segmentul de HVS-am haplotipuri. Prin urmare, după câteva mii de ani, toți s-ar putea să fi suferit mutații de mai multe ori.
Pe de altă parte, este mai mic numărul de tastat markeri cea mai mare probabilitatea de identice haplotipuri ca urmare a homoplasy sau de un vechi stramos comun., Cu alte cuvinte, adoptarea haplotipurilor modale ca markeri pentru anumite evenimente istorice trebuie să țină seama de rezoluția temporală oferită de numărul de variante incluse în analize. Haplotipurile y definite de zeci de variante minimizează riscul de identitate în funcție de stat (IBS) sau convergență, permițând astfel o putere de diagnosticare ridicată la cheltuielile unei rezoluții temporale superficiale (TMRCA recent). Haplotipuri definite de 6 sau 12 Str, cum ar fi CMH și LMH sau eCMH și eLMH, permite o mai profundă rezoluție temporală, dar cu o creștere de eroare din cauza mutații recurente., Adoptarea secvențelor mtDNA întregi ar putea îmbunătăți rezoluția anumitor filogenii materne. Cu toate acestea, conform loci-ului many-Y-STR, acest lucru ar afecta în mod necesar compromisul dintre putere pentru a discrimina între IBD și IBS și rezoluția temporală.
rezultatele noastre sunt o demonstrație a acestui argument: utilizarea motivelor haplotipice non-recombinante ca markeri de diagnostic ai strămoșilor evrei s-a dovedit a fi puternic înșelătoare atunci când nu este susținută de cunoștințe în regiuni mai informative sau secvențe întregi., Dintre motivele presupuse anterior pentru a urmări strămoșii evrei specifici, niciunul nu a rezultat identic prin descendență (IBD), care este moștenit fără modificări de la un strămoș comun. Cu puține excepții, motivele, fie din NRY, fie din mtDNA, au fost observate în cel puțin două linii independente, uneori aparținând unor grupuri etnice cu afilieri culturale sau geografice diferite., Pentru a explica polyphyletic model de haplotip distribuția pe loturi analizate, avem în vedere, ca cel mai succintă explicație, mai multe fondator evenimente și/sau amestecare a genomice piscine printr-o lungă istorie de dispersie și de amestec de comunități Evreiești din temelia lor.pentru a face un exemplu, semnătura „CMH”, în versiunea sa clasică și extinsă, a fost observată în multe haplotipuri de locuitori din Peninsula Arabică cu nume tipice arabe, precum și în multe persoane evreiești aparținând haplogrupurilor J1 și J2., Distribuția de CMHs de etnie și haplogroups este de natură să sugereze că gena conversii, adopțiile și nelegitim paternities ar putea afecta doar marginal rezultatele excepția cazului în care acestea au fost multiple și în principal a avut loc cu sute de ani în urmă.
o explicație mai ușoară este că, între 7600 și 10.400 de ani bp (IÎ 95%), „haplotipul Modal Cohen” a fost un haplotip ancestral pentru locuitorii istorici ai Peninsulei Arabe., În urmă cu aproximativ 4000 ± 520 de ani, populația evreiască stabilită a purtat acest „haplotip modal” împreună cu viitorii arabi, care la acea vreme aveau un strămoș comun cu viitorii evrei (Klyosov, 2010). În jurul secolului al X-lea d.hr., o „CMH recentă” ușor modificată s-a despărțit de „CMH mai vechi” (în formate de haplotip mai extinse), în timp ce ambele conțineau semnătura markerului 6 a „CMH”, care este încă prezentă în multe haplotipuri Arabe. Acest „CMH recent” a devenit haplotipul ancestral pentru o linie evreiască separată, deși recentă, în haplogrupul J1., Dacă luăm în considerare doar haplotipurile” CMH ” din această populație, poate fi identificat un strămoș comun care a trăit în urmă cu aproximativ 1255-986 de ani.
Concentrându-se pe mitocondriale motive, prezența de variante la rapid mutații site-uri, cum ar fi 16,311 și 16,519 crește potențialul pentru apariția de mutații recurente la HVS-am segmentele. Acest lucru este deosebit de important în haplogroup K., Monophyly și recente TMRCAs, și anume genealogii trasabile la un singur strămoș recent, ar putea fi invocate pentru K1, K2, și N motive, în timp ce amestec și mai multe fondatorii ar trebui să fie invocate pentru K3 motiv, în lipsa de o mai bună rezoluție, în conformitate cu completă a genomului și re-secventiere de date.un element suplimentar de incertitudine este jucat de alegerea ratei mutaționale., Haplotipul mutații schimbarea alele lor imprevizibil, și doar un număr mediu de mutații peste un timp dat poate fi anticipat cu o anumită probabilitate, bazat pe rata de mutație constante și pe cât de „vechi” este grupul de haplotipuri în termeni de o durata de timp de la strămoșul lor comun.
problema care este cea mai potrivită constantă a ratei de mutație a haplotipului care trebuie aplicată pentru urmărirea căilor istorice ale populațiilor umane a fost dezbătută fierbinte (vezi Soares et al., 2009; Wei și colab., 2013) și disponibilitatea recentă a datelor genomului întreg și a resecvenței nu au rezolvat-o., Este larg acceptat faptul că numărul de mutații între haplotipurile divergente se saturează destul de repede din cauza mutațiilor recurente, în special la markerii STR. De asemenea, este implicit că ratele calibrate pe modele de ramificare infinite și pe intervale de timp evolutive umflă TMRCAs de haplotipuri care au ajuns să se abată în vremurile istorice. Este cazul ratei propuse de Zhivotovsky și colab. (2004), care a fost folosit de Ciocan și colegii să susțină că vârsta de eCMHs este compatibil cu fundația de Cohanim preoției (vezi critica în Klyosov, 2009b)., De asemenea, este evident că estimările bazate pe germeni, tați-fii sau pedigree cu înrădăcinare profundă sunt adesea susținute de statistici foarte slabe, mutațiile observate la un anumit marker fiind foarte puține sau deloc.pentru a complica imaginea este conceptul că o rată constantă nu există în lumea reală. În sine, Numărul de ani în generație este o valoare plutitoare, depinde de obiceiurile culturale, opiniile religioase, vârsta la nașterea copilului, nutriția, sănătatea și alte condiții de viață pe un anumit teritoriu la un moment dat., Mai mult, probabilitatea apariției unei mutații noi depinde de structura regiunii genomice în care se întâmplă, iar soarta acesteia depinde în mare măsură de dimensiunea și demografia comunității din care face parte.de regulă ,cu cât timpul este mai lung pentru un strămoș comun al unui grup de haplotipuri, cu atât mai puțin „diagnosticarea” unui motiv și estimările de timp mai incerte., Numai grupuri cu ultimii strămoși comuni au destul de previzibil motive, cum este cazul de Evrei Ashkenazi la unele mtDNA haplogroups și Y-R1a sub-tipurile extins în ultimele mii de ani, pentru că nu a mai rămas mult timp, deoarece strămoș comun, și lui haplotip este încă în jurul valorii de a avea relativ puține mutații.toate motivele propuse s-au dovedit a fi distribuite inegal între indivizi grupați în funcție de strămoșii lor evrei și nu evrei auto-raportați, aproape întotdeauna cu o îmbogățire semnificativă în Evrei (tabelele 4, 5)., Cu toate acestea, compoziția eterogenă a haplotipurilor care le conțin a afectat orice încercare de a-și întâlni corect originea. Tastarea SNP cu rezoluție mai mare și, sperăm, disponibilitatea secvențelor complete, ar putea ajuta la rafinarea filogeniei unor astfel de markeri, clarificând în cele din urmă rolul și timpul lor de la înființarea grupurilor evreiești.,
În concluzie, în timp ce distribuția observată a sub-tipurile de haplotipuri la mitocondriale și cromozomul Y non-recombinant genomul ar putea fi compatibil cu fondatorul evenimente în ultimii ori la originea unor grupuri de Evrei ca Cohenite, Levitul, Ashkenazite, de ansamblu substanțiale polyphyletism precum și sistematică apariția în non-grupuri de Evrei subliniază lipsa de sprijin pentru a le utiliza fie ca markeri de origine Evreiască sau povești Biblice.,
Declarație privind conflictul de interese
autorii declară că cercetarea a fost efectuată în absența oricăror relații comerciale sau financiare care ar putea fi interpretate ca un potențial conflict de interese.
confirmare
autorii ar dori să recunoască recenzenții pentru comentariile lor critice și stimulative.