visão geral
ácido gordo β-oxidação é um processo em várias etapas pelo qual os ácidos gordos são decompostos por vários tecidos para produzir energia. Os ácidos gordos entram principalmente numa célula através de transportadores de proteínas de ácidos gordos na superfície celular . Os transportadores de ácidos gordos incluem a translocase de ácidos gordos (FAT/CD36), as proteínas de transporte de ácidos gordos específicas dos tecidos (FATP) e a proteína de ligação de ácidos gordos ligada à membrana plasmática (FABPpm) . Uma vez dentro da célula, um grupo CoA é adicionado ao ácido gordo por acil-CoA sintase gorda (FACS), formando acil-CoA de cadeia longa., Carnitina palmitoiltransferase 1 (CPT1) a conversão da acil-CoA de cadeia longa em acilcarnitina de cadeia longa permite que a fracção de ácidos gordos seja transportada através da membrana mitocondrial interna através da carnitina translocase (CAT), que troca acilcarnitinas de cadeia longa por carnitina. Uma membrana mitocondrial interna CPT2 converte então a acilcarnitina de cadeia longa para acil-CoA de cadeia longa. A acil-CoA de cadeia longa entra na Via de β-oxidação de ácidos gordos, o que resulta na produção de um acetil-CoA a partir de cada ciclo de β-oxidação de ácidos gordos., Este acetil-CoA entra então no ciclo do ácido tricarboxílico mitocondrial (TCA). O NADH e o FADH2 produzidos tanto pela oxidação β de ácidos graxos quanto pelo ciclo TCA são usados pela cadeia de transporte de elétrons para produzir ATP. A Figura 1 apresenta uma panorâmica da oxidação de ácidos gordos.
a Figura 1. Oxidação de ácidos graxos é o processo pelo qual os ácidos graxos são decompostos para produzir energia. Os ácidos gordos entram principalmente numa célula através de transportadores de proteínas de ácidos gordos na superfície celular., Uma vez lá dentro, o FACS adiciona um grupo CoA ao ácido gordo. A CPT1 converte então a acil-CoA de cadeia longa em acilcarnitina de cadeia longa. A fracção de ácidos gordos é transportada pelo gato através da membrana mitocondrial interna. A CPT2 converte então a acilcarnitina de cadeia longa para acil-CoA de cadeia longa. A acil-CoA de cadeia longa pode então entrar na Via de β-oxidação do ácido gordo, resultando na produção de um acetil-CoA de cada ciclo de β-oxidação. Este acetil-CoA, em seguida, entra no ciclo TCA., O NADH e o FADH2 produzidos tanto pela β-oxidação quanto pelo ciclo TCA são usados pela cadeia de transporte de elétrons para produzir ATP.

Papel de Ácido Graxo de Abastecimento na Regulação Ácido Graxo β-Oxidação
Celular ácido graxo de transporte:
houve um esforço considerável nos últimos anos para elucidar os mecanismos pelos quais os ácidos graxos são absorvidos pelas células, particularmente determinar se os ácidos graxos são transportados através da membrana celular por difusão simples ou se este transporte é facilitado pela membrana proteínas associadas., Embora vários resultados apoiem ambos os métodos de transporte, acredita-se que o transporte por proteínas associadas à membrana seja o meio predominante de absorção de ácidos gordos nas células . Foram identificadas várias proteínas de membrana que facilitam a captação de ácidos gordos celulares. Os transportadores de ácidos gordos associados à membrana CD36/FAT, FABPpm e FATPs diferem no seu peso molecular e grau de modificação pós-translacional . Stremmel et al. relataram que os anticorpos dirigidos contra o FABPpm do fígado de rato inibem a absorção de ácidos gordos por hepatócitos, adipócitos e cardiomiócitos em 50-75%., Este resultado sugere que uma porção significativa da captação de ácidos gordos depende do transporte mediado pelas proteínas em diferentes tipos de células .Schaffer e Lodish descobriram o Lipp por uso de uma estratégia de clonagem de expressão em adipócitos . A expressão desta proteína integral de membrana de 63 kDa numa linha celular estável de fibroblastos resultou num aumento de 3 a 4 vezes no transporte de ácidos gordos de cadeia longa. A identificação deste primeiro GAFI levou à descoberta de várias outras isoformas do GAFI (GAFI-1-6) . O LIPP1 é predominantemente expresso nos músculos do coração e do esqueleto ., O FATP2 e o FATP5 são expressos principalmente no fígado, onde estão envolvidos no metabolismo lipídico hepático em associação com acil-CoA sintetase. O FATP4 é essencial para a absorção de lípidos dietéticos e tem um papel fundamental na estrutura e função da pele normal. Até à data, o FATP3 e o FATP6 demonstraram ter pouca ou nenhuma função de transporte de ácidos gordos .o papel da proteína de membrana CD36 / gordura na absorção de ácidos gordos e β-oxidação nos mamíferos foi amplamente estudado., CD36 / FAT é uma proteína de membrana translocase de 88 kDa expressa em proporção à taxa de oxidação de ácidos gordos no tecido muscular (por exemplo, é expressa mais no coração do que no músculo esquelético) . CD36 / gordura está envolvida na angiogênese e inflamação, bem como metabolismo lipídico. A transferência de fibroblastos com CD36/gordura resultou num aumento das taxas de absorção de ácidos gordos . Ao contrário do FATP, o CD36 / gordura tem a capacidade de translocação entre os endossomas intracelulares e a membrana plasmática das células, o que permite que o CD36/gordura desempenhe um papel crítico na regulação da captação de ácidos gordos., A contracção da insulina e do músculo pode estimular a translocação CD36/gordura das reservas intracelulares para a membrana plasmática, o que leva a uma maior captação e β-oxidação de ácidos gordos. A activação da FoxO1 pode levar à translocação CD36/gordura, o que resulta num aumento da oxidação β de ácidos gordos, juntamente com a acumulação de triacilglicerol . Isto sugere um papel significativo da sinalização intracelular na função CD36/FAT.o mecanismo exacto de indução da translocação CD36/gordura ainda é Desconhecido., No entanto, assume-se que a activação da proteína cinase activada pela AMP (AMPK) e o estado energético das células musculares podem participar na resposta de translocação CD36/gordura . Além disso, a estimulação da proteína cinase c (PKC) nos cardiomiócitos induz a translocação CD36/gordura, indicando um possível papel do cálcio no processo de translocação . Por outro lado, a inibição do receptor de sinalização extracelular kinase (ERK) pode bloquear a translocação CD36/gordura induzida pela contracção muscular ., A modificação pós-translacional do CD36 / gordura através da ubiquitinação também pode regular os níveis de proteína intracelular do CD36 / gordura, visando a proteína para degradação. Assim, a insulina aumenta a disponibilidade de CD36/gordura para translocação por inibição da ubiquitinação. No entanto, os ácidos gordos promovem a ubiquitinação, o que leva à degradação CD36/gordura .esterificação de ácidos gordos em acil-CoA: um ácido gordo deve ser convertido em acil-CoA gordos para entrar na mitocôndria e ser oxidado ., A enzima responsável pela esterificação dos ácidos gordos em acil-CoA gordos de cadeia longa é a FACS. Para esta reação, a FACS consome o equivalente a dois ATP. Outra enzima, tioesterase citosólica (CTE), pode remover a CoA convertendo a acil-CoA gorda de volta para um ácido gordo. A acil-CoA gorda pode ser convertida em acil-carnitina, permitindo que seja transportada para as mitocôndrias e entrar na oxidação do ácido gordo Por β ou convertida em metabolitos lipídicos (triacilglicerol, diacilglicerol, ceramida, etc.).,o acetil-CoA carboxilase, malonil-CoA descarboxilase, eixo malonil-CoA:
Acetil-CoA carboxilase (ACC) é uma enzima central envolvida na biossíntese de ácidos gordos e β-oxidação de ácidos gordos. A ACC catalisa a carboxilação da acetil-CoA produzindo malonil-CoA, que pode ser usada pela ácido graxo sintase para a biossíntese de ácidos graxos . Enquanto o malonil-CoA é usado como substrato para a biossíntese de ácidos gordos, o malonil-CoA é também um potente inibidor da captação de ácido gordo mitocondrial secundário à inibição do CPT1 (Figura 2) ., Existem duas formas de ACC, uma isoforma 265 kDa ACC1, que é altamente expressa no fígado e tecido adiposo, e uma isoforma de 280 kDa ACC2, que é mais específica para órgãos altamente metabólicos, como o músculo esquelético e o coração . O AMPK desempenha um papel importante na regulação ACC1 e ACC2 por fosforilação e inibição da atividade ACC. Em situações de maior demanda de energia, o AMPK é ativado, onde então fosforila e inativa ambas as isoformas de ACC (Figura 3). A inibição de ACC2 pode levar a um aumento na oxidação β de ácidos graxos, enquanto a biossíntese de ácidos graxos diminui quando o ACC1 é inibido .,
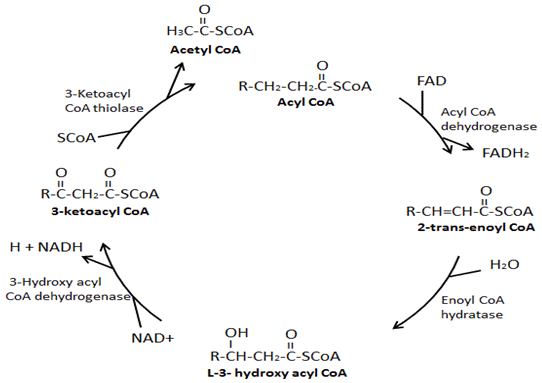
Figura 2. A via de β-oxidação do ácido gordo
As quatro principais enzimas envolvidas Na β-oxidação são: acil-CoA desidrogenase, enoil-CoA hydratase, hidroxi acil-CoA desidrogenase e cetoacil-CoA tiolase. A acil-CoA desidrogenase cria uma ligação dupla entre o segundo e terceiro carbonos abaixo do grupo CoA em acil-CoA e no processo produz um FADH2., Em seguida, a enoil-CoA hydratase remove a dupla ligação formada, no processo de adição de um grupo hidroxila ao terceiro carbono para baixo do grupo CoA e um hidrogênio no segundo carbono para baixo do grupo CoA. Hydroxyacil-CoA desidrogenase remove o hidrogênio no grupo hidroxilo apenas ligado e no processo produz um NADH. No passo final, a cetoacil-CoA tiolase Liga um grupo CoA ao terceiro carbono para baixo a partir do grupo CoA resultando na formação de duas moléculas, uma acetil-CoA e uma acil-CoA que é dois carbonos mais curto.,
a regulação a longo prazo da ACC depende da regulação da sua expressão genética. Vários fatores transcritionais podem regular a expressão do gene ACC, incluindo proteína de ligação do elemento regulador do esterol (SREBP1a e SREBP1c) e proteína de ligação do elemento de resposta dos hidratos de carbono (ChREBP) . SREBP é regulado pela insulina, que promove o retículo endoplasmático srebp1c a ser clivado e translocado para o núcleo, levando à estimulação da expressão ACC., Além disso, os fatores de transcrição peroxissoma proliferator activated receptor γ proliferação 1 (PGC-1) α e β podem estimular a expressão de SREBP1a e SREBP1c, ambos os quais têm um papel vital na lipogênese. A expressão de ChREBP pode ser induzida por altas concentrações de glicose, resultando no ChREBP ativado promovendo a expressão de ACC1 e ácido graxo sintase . O fator respiratório Nuclear-1 (NRF-1) é um modulador principal da expressão proteica mitocondrial e da biogênese mitocondrial, ambos importantes para uma maior capacidade de oxidação de ácido gordo mitocondrial ., Por exemplo, Adam et al. demonstrou que a sobreexpressão NRF-1 resulta na inibição da actividade do promotor do gene ACC2 no coração dos mamíferos, o que aumenta as taxas de β-oxidação do ácido gordo mitocondrial, promovendo assim a capacidade energética intracelular global .
malonil-CoA descarboxilase (MCD) é a enzima responsável pela descarboxilação do malonil-CoA ao acetil-CoA . Geralmente, o nível de malonil-CoA é diminuído quando a atividade MCD é aumentada, resultando em uma taxa elevada de oxidação de ácidos graxos., Tem sido relatado que cinases proteicas que fosforilato e inibir ACC podem ativar MCD . No entanto, a MCD parece ser principalmente regulada por meios transcritionais (discutidos mais tarde). Portanto, MCD e ACC parecem trabalhar em harmonia para regular o pool de malonil-CoA que pode inibir o CPT1 .
mitocondrial carnitina palmitoyl transferase (CPT):
a isoforma CPT, CPT1, reside na superfície interna da membrana mitocondrial exterior e é um dos principais locais de regulação da captação de ácidos gordos mitocondriais ., Como mencionado, a CPT1 é potentemente inibida pela malonil-CoA, o produto da ACC que se liga ao lado citosólico da CPT1. Os mamíferos expressam três isoformas de CPT1, que são codificadas por genes diferentes. A isoforma hepática (CPT1a), a isoforma muscular (CPT1ß), e uma terceira isoforma de CPT1 (CPT1c), que é principalmente expressa no cérebro e nos testículos . Mais especificamente, o coração expressa duas isoformas de CPT1, uma isoforma de 82 KDa( CPT1a) e a isoforma predominante de 88 KDa (CPT1ß) (que tem a maior sensibilidade à inibição de malonil-CoA)., A insulina e a hormona tiroideia podem regular a sensibilidade da CPT1a no fígado; no entanto, a isoforma CPT1ß não é afectada . Estudos anteriores relataram que os níveis de malonil-CoA estão inversamente correlacionados com as taxas de β-oxidação de ácidos gordos . Além disso, estudos em ratos nocaute ACC2 sugerem duas piscinas celulares de malonil-CoA, malonil-CoA produzidas pela ACC1 (utilizadas principalmente para a lipogénese), e um pool citosólico de malonil-CoA produzido pela ACC2 envolvido na regulação da CPT1 e Da β-oxidação de ácidos gordos .,
ácido gordo β-oxidação mitocondrial
a via de β-oxidação do ácido gordo:
ácido gordo β-oxidação é o processo de decomposição de uma molécula de acil-CoA de cadeia longa em moléculas de acetil-CoA. O número de acetil-CoA produzido depende do comprimento de carbono do ácido gordo sendo oxidado. Este processo envolve uma variedade de enzimas, com as quatro principais enzimas envolvidas Na β-oxidação de ácidos graxos sendo, por ordem, acil-CoA desidrogenase, enoil-CoA hydratase, hidroxiacil-CoA desidrogenase e cetoacil-CoA tiolase (Figura 3) ., No final de cada ciclo de β-oxidação, duas novas moléculas são formadas, uma acetil-CoA e uma acil-CoA que é dois carbonos menores. Além disso, durante a β-oxidação NADH e FADH2 são formados. Um FADH2 é produzido durante a reação catalisada pela acil-CoA desidrogenase. Um NADH é produzido durante a reação catalisada pela hidroxiacil-CoA desidrogenase. O FADH2 e o NADH produzidos durante o processo de oxidação Por β de ácidos graxos são usados pela cadeia de transporte de elétrons para produzir ATP., Existem isoformas diferentes destas enzimas de β-oxidação, que têm afinidades diferentes para diferentes comprimentos das cadeias de ácidos gordos. Por exemplo, há uma acil-CoA desidrogenase de cadeia muito longa, uma acil-CoA desidrogenase de cadeia longa, uma acil-CoA de cadeia média desidrogenase, e uma acil-CoA de cadeia curta desidrogenase. Curiosamente, as isoformas enoil-CoA hydratase, hidroxiacil-CoA desidrogenase e cetoacil-CoA específicas para ácidos gordos de cadeia longa formam um complexo enzimático na membrana mitocondrial interna.
a Figura 3., Os principais locais de regulação Da β-oxidação de ácidos gordos
ácido gordo β-oxidação é regulada em níveis múltiplos. Esta figura mostra algumas das formas como a oxidação de ácidos gordos β é regulada. 1. A regulação pode ocorrer ao nível da entrada de ácidos gordos na célula. AMPK, PKC e PPARy regulam positivamente a atividade do CD36 / FATP. 2. A regulação também ocorre através da regulação dos níveis de acetil-CoA e malonil-CoA. O AMPK inibe a ACC, resultando num aumento dos níveis de acetil-CoA/numa diminuição dos níveis de malonil-CoA e num aumento da oxidação dos ácidos gordos. O malonil-CoA inibe a oxidação dos ácidos gordos inibindo o CPT1., 3. A regulamentação transcritional também está envolvida na regulação da oxidação de ácidos gordos Por β. PGC-1α, um coregulador de fator de transcrição, e o Fator de transcrição PPARa agem no núcleo para aumentar a transcrição de genes mitocondriais, genes de utilização de ácidos graxos, e outros fatores de transcrição.são necessárias enzimas auxiliares para a oxidação β de ácidos gordos insaturados e ácidos gordos de cadeia ímpar. Os ácidos gordos ímpares são decompostos por β-oxidação em moléculas de acetil-CoA e propionil-CoA., Embora o propionil-CoA possa ser metabolizado através de vias alternativas, é principalmente metabolizado na célula para succinil-CoA por três enzimas (carboxilase propionil-CoA, epimerase metilmalonil-CoA e mutase metilmalonil-CoA) . Esta succinil-CoA pode então entrar no ciclo TCA. Em comparação com ácidos gordos pares, ácidos gordos ímpares ocorrem pouco frequentemente na natureza . As duas enzimas auxiliares, a isomerase enoil-CoA e a redutase 2,4-dienoil-CoA são necessárias para a oxidação completa dos ácidos gordos insaturados ., Durante a β-oxidação, o ciclo em que o cis-ligação dupla começa no terceiro carbono do acyl-CoA, o primeiro passo envolve a enoyl-CoA isomerase isomerizing antes enoyl-CoA hydratase, e as outras duas enzimas, pode agir sobre o acyl-CoA. Uma ligação dupla num carbono par-numerado requer ambas as enzimas auxiliares. Uma vez que a ligação dupla está no quarto carbono do acil-CoA no início de um ciclo de β-oxidação, ele começa a ser oxidado. Após a acção da acil-CoA desidrogenase, a 2,4-dienoil CoA redutase actua na acil-CoA, seguida da enoil-CoA isomerase., Enoil-CoA hydratase então age sobre acil-CoA e o processo retoma sua ordem normal.controlo alostérico Da β-oxidação de ácidos gordos: a actividade das enzimas de β-oxidação de ácidos gordos é afectada pelo nível dos produtos das suas reacções . Cada uma das enzimas de β-oxidação é inibida pelo intermediário acil-CoA específico que produz . Curiosamente, 3-cetoacil-CoA também pode inibir a enoil-CoA hydratase e acil-CoA desidrogenase . a β-oxidação também pode ser allostericamente regulada pela razão NADH/NAD+ e nível acetil-CoA / CoA., Um aumento das razões NADH/NAD+ ou acetil-CoA / CoA resulta na inibição da oxidação por ácidos gordos. Os aumentos na razão acetil-CoA/CoA demonstraram, especificamente, conduzir à inibição do feedback da cetoacil-CoA tiolase .a β-oxidação de ácidos gordos também pode ocorrer em peroxissomas. Em animais, acredita-se que os peroxissomas sejam importantes na decomposição inicial de ácidos gordos de cadeia muito longa e ácidos gordos ramificados de metilo . As enzimas envolvidas na oxidação de ácidos gordos em peroxissomas são diferentes das mitocôndrias., Uma diferença importante é a acil-CoA oxidase, a primeira enzima na oxidação β-peroxisoma, que transfere o hidrogênio para o oxigênio produzindo H2O2 em vez de produzir FADH2. O H2O2 é dividido em água por catalase. Importante, os intermediários de acilo-CoA gordos formados durante a oxidação-β são os mesmos em peroxissomas e mitocôndrias. Os peroxissomas também contêm as enzimas necessárias para a α-oxidação, que são necessárias para a oxidação de alguns ácidos gordos com ramos de metilo.,as proteínas envolvidas na oxidação de ácidos gordos Por β são reguladas por mecanismos tanto de transcrição como de pós-transcrição. Há uma série de fatores de transcrição que regulam a expressão dessas proteínas. Os receptores activados pelo proliferador de peroxissomas (PPARs) e um co-activador do factor de transcrição PGC-1α são os reguladores de transcrição mais conhecidos da β-oxidação de ácidos gordos . PPARs e retinóide X receptor heterodimerize e se ligue a promotores de genes contendo o elemento de resposta PPAR ., Exemplos de proteínas envolvidas em ácidos graxos essenciais β-oxidação, que são transcriptionally regulamentada pelo PPARs incluem FATP, acyl-CoA sintetase (ACS), CD36/FAT, MCD, CPT1, long-chain acyl-CoA desidrogenase (LCAD), e médio-chain acyl-CoA desidrogenase (MCAD) . O receptor α (ERRa) relacionado ao estrogênio também tem sido implicado na regulação da oxidação do ácido gordo β, tendo sido demonstrado que também regula a transcrição do gene que codifica MCAD . Ligantes que se ligam e modulam a atividade de PPARa, δ e γ incluem ácidos graxos .,
os genes regulados por cada um dos PPARs variam entre os tipos de tecidos. Por exemplo, o músculo esquelético PPARδ, mas não PPARa, eleva a expressão de CPT1 . As isoformas do PPAR também são expressas diferentemente entre os tipos de tecidos . Enquanto a proteína PPARδ tende a ser ubiquitosamente expressa, a PPARa é predominantemente expressa em tecidos altamente metabólicos (ou seja, coração, músculo esquelético e fígado) e o PPARy é predominantemente expresso em tecidos como tecido adiposo . Até recentemente, não acreditava-se que o PPARy desempenhasse um papel significativo na regulação da oxidação de ácidos graxos β., No entanto, estudos recentes de nocaute e sobreexpressão sugeriram que o PPARy pode ter um papel na regulação da oxidação de ácidos graxos β. Over expressing PPARy in cardiac muscle results in increased mRNA levels for fatty acid β-oxidation proteins .
O co-activador transcritional PGC-1α liga-se e aumenta a actividade dos PPARs e ERRa para regular a β-oxidação de ácidos gordos . PGC-1α modula a atividade de uma série de fatores de transcrição que podem aumentar a expressão de proteínas envolvidas na oxidação de ácidos graxos β, o ciclo TCA e a cadeia de transporte de elétrons., Por exemplo, o aumento da expressão proteica PGC-1α induz uma massiva biogénese mitocondrial no músculo esquelético .
PGC-1α é regulado tanto ao nível do gene como ao nível das proteínas. AMPK aumenta a atividade da proteína PGC-1α pré-existente através de dois mecanismos propostos. A primeira é a fosforilação de PGC-1α em resíduos de treonina e serina, resultando num aumento global da actividade PGC-1α . AMPK também pode aumentar a atividade de PGC-1α ativando sirtuin 1 (SIRT1). SIRT1 pode então deacetilato PGC-1α, aumentando sua atividade ., Acredita-se que o AMPK aumenta os níveis de mRNA PGC-1α regulando a ligação dos factores de transcrição a sequências específicas localizadas no promotor do gene PGC-1α, que incluem dois locais de MEF, um local do elemento de resposta do campo (CRE) e a região de GATA/Ebox . AMPK regula os locais do MEF por fosforilação do GEF ,uma proteína que pode mediar o movimento do MEF2 para o núcleo. O AMPK pode aumentar a ligação ao local CRE por fosforilação da proteína de ligação do elemento de resposta do campo (CREB) 1 e de outros membros da família CREB que se ligam às regiões promotoras do CRE ., Como outro exemplo, ácidos graxos livres também podem regular a expressão de proteína PGC-1α. Por exemplo, uma dieta rica em gorduras pode elevar os níveis de PGC-1α no músculo esquelético do rato .
conclusões
a β-oxidação de ácido gordo é a principal via metabólica que é responsável pela discriminação mitocondrial da acil-CoA de cadeia longa à acetil-CoA. Este processo envolve muitos passos que são regulados no nível transcritional e pós-transcritional., A regulação transcritional envolve PPARs, SREBP1 e PGC-1α, enquanto o nível pós-transcriptional envolve principalmente o controle alostérico da oxidação de ácidos graxos β, bem como a regulação ACC, MCD e CPT. Ambos os mecanismos funcionam em harmonia para garantir um fornecimento contínuo de acil-CoA de longa cadeia para a β-oxidação, e produtos de β-oxidação para a produção de energia mitocondrial.
agradecimentos: GDL é um cientista da fundação do património de Alberta para a investigação médica